为什么虚假信息传播快?我们能预测攻击性行为吗?
为什么虚假消息
会在网络上被疯狂转发?
文章来源:神经现实
在信息爆炸的今天,各种新闻头条往往真假难辨。有些明明看着就是捏造的标题党,为啥还是会在网络上大行其道,屡禁不绝?我们作为普通人,又如何避免被标题蒙蔽?最近发表在《自然》的一项研究发现了一个有趣的现象:人们转发一则消息,并不代表他们相信消息的真实性。
作者提出了三个假说来解释为什么人们会转发虚假消息:(1)混淆了正确或错误;(2)注重其他因素(比如个人喜好,党派倾向)大于准确性;(3)没有对消息的正确性产生足够的注意。到底是哪种原因解释了虚假消息的传播呢?
为了探究第一个假说的可能性,研究者招募了1000多名受试者,并给他们看36条消息(包括标题,内容总结句,和图片),这36条消息中有一半为准确的,一半为虚假的;一半更倾向于民主党,另一半则更倾向于共和党。受试者被随机分配去单纯判断这些消息的真实性(准确状态,accuracy condition),或是单纯决定自己是否会考虑转发(分享状态,sharing condition)。结果他们发现,在准确状态下,受试者都能很好的分辨标题的真假。有趣的是,如果是在分享状态,标题是否符合受试者的政治立场,对他们是否分享消息的决定影响更大(远大于消息准确性的影响)。也就是说他们不论准确与否,都更愿意分享与自己立场一致的消息——这一发现也与实际twitter用户分享的行为一致。
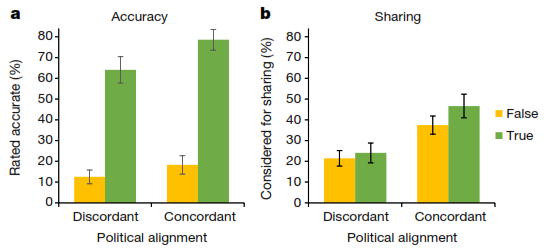 - Pennycook et al。, Nature-
- Pennycook et al。, Nature-那么,是不是说相比于消息的准确性,人们更重视个人喜好和政治倾向呢?研究者随后设计了调查问卷,让受试者给消息的各种考量因素的重要性评分。调查结果显示,绝大部分受试者都认为,相比于“政治立场一致”,“有趣”,“令人惊讶”等因素,“消息的准确性”是才是更重要的。所以第二个假说也并不成立。
还有一种可能性是,现在的社交媒体的标题经常会引导读者注意到消息的其它方面,以此捕捉他们眼球,取悦读者,或者凸显读者所在的组织团体身份,所以常常导致读者忽略消息的准确性。为了弄清楚这个问题,研究者想了个办法巧妙地引导受试者在测试前关注到准确性(即准确性的启动效应,priming effect):如果受试者只关心消息是否符合个人三观的话,那么准确性的启动效应则不应该对消息的转发行为产生影响。于是,在接下来的测试中,受试者被要求先要对一个没有政治倾向的消息的准确性评分,然后再完成与之前相同的测试;或者更严格的,在决定是否转发每一条消息之前,受试者都要先给该消息的准确性评分。结果发现,当准确性的启动效应开启之后,大家转发虚假消息的频率大大减小了,尤其是对于与个人三观一致但虚假的消息,被转发的几率大大降低。这些实验说明,对消息没有足够注意是导致转发虚假原因的主要原因。
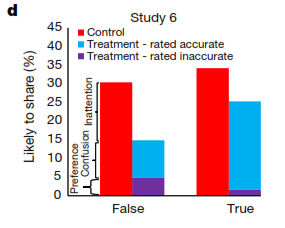 - Gordon Pennycook et al。, Nature-
- Gordon Pennycook et al。, Nature-最后小编要敲黑板划重点了:作为一个理性的读者,下次看到了令人拍案叫绝,血脉喷张的标题,在按下转发按钮之前,记得先问问自己消息准确不准确哦!
新事物的价值编码
原来藏在网格里·····
在对非人灵长类动物的价值决策的大多数研究中,实验范式通常包括长时间的训练阶段:动物总是在训练中先将特定刺激和价值建立联系,然后在测试中通过衡量刺激相对应的价值来进行决策;而在真实世界中的决策并非如此——我们常常面临着从未见过的选项,因此需要根据已知的经验去推断某个陌生的对象的价值。要完成这一过程,就要求大脑对新事物的价值的表征不能完全基于特定的选项(option-specific representation);大脑需要从新选项中提取出某些特征,然后根据经验中出现过的、具有相同特征选项的价值来评估新选项的价值。通常引导价值推断的特征不止一个维度,大脑需要在价值空间内对多维度价值信息进行整合,从而做出最优决策。
Bongioanni等人设计了三个实验来探究清醒猕猴是否能够对新对象进行价值推断,以及这一过程背后不同于常规决策过程的神经机制。实验所采用的数据集设计如Fig.1a&1b所示。引导猕猴进行价值推断的维度有两个,第一个维度代表着奖励的大小(magnitude,即完成任务后获得果汁的滴数),第二个维度代表着获得奖励的概率(probability)。
实验包括两个训练集(Fig.1a上、中):第一个训练集保持获得奖励的概率为0.6不变,通过变化刺激的颜色来表示从1-10不等大小的奖励;第二个训练集保持奖励大小为5不变,通过变化刺激的点数来表示从0.1-1.0不等的获得奖励的概率。研究者采用这两个训练集对猕猴进行了3个月的训练。
实验的测试集则在两个维度上变化形成了6种(2ⅹ3)不同的条件(Fig.1a下):在第一个维度上,两种刺激可能是熟悉的(familiar,训练集中出现过)或陌生的(novel,训练集中未出现过);在第二个维度上,两种刺激可能是价值指向统一的(consistent,同一个选项的获得奖励概率和奖励大小都更优)、一维变化的(1-dimension,两个选项只在获得奖励概率和奖励大小中的一个维度有差异)或者价值指向不统一的(inconsistent,某一个选项的获得奖励概率更高,另一个选项的奖励更大)。
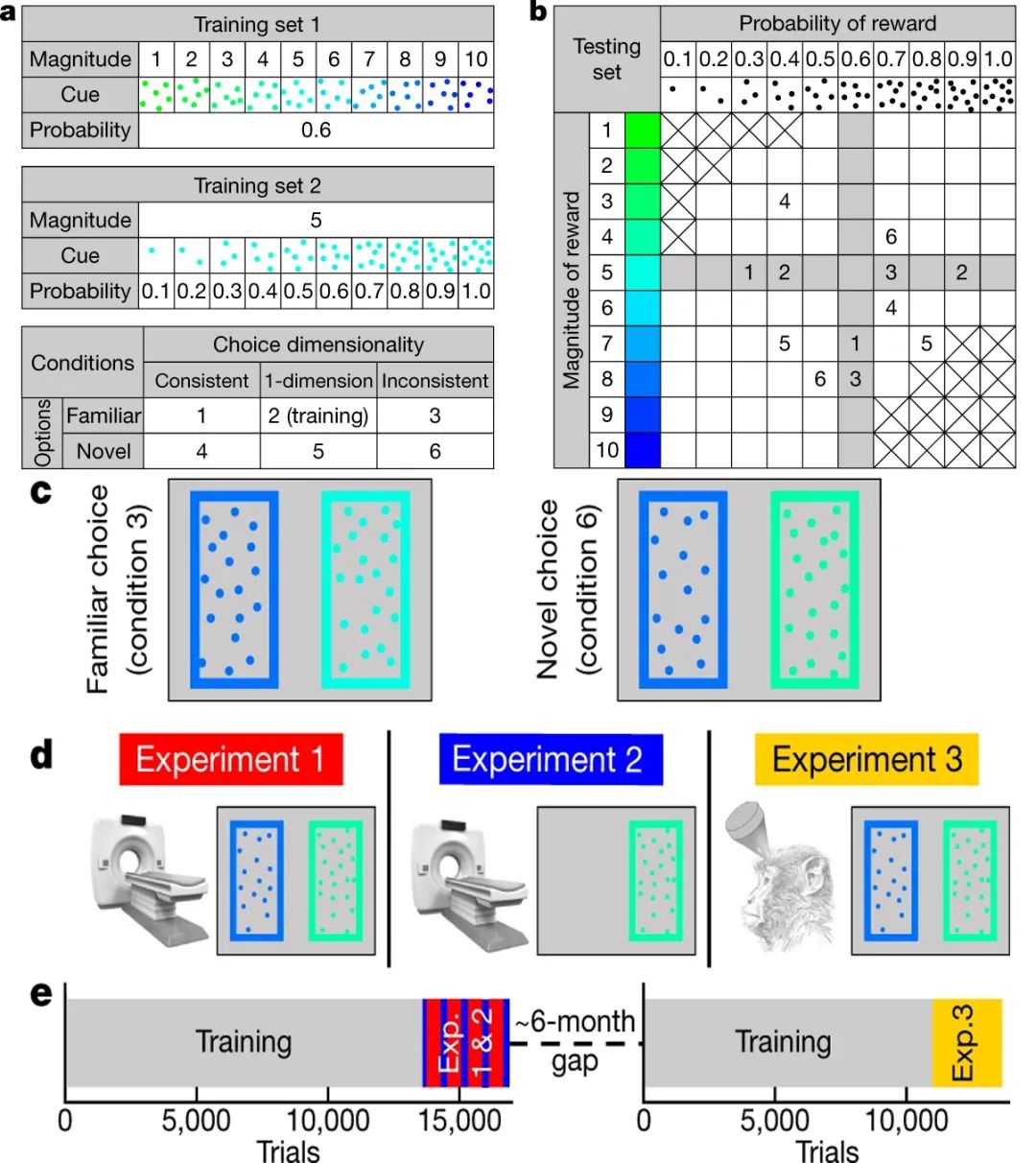
Fig.1|实验设计
(a)上:训练集1,刺激所代表获得奖励的概率设为0.6固定不变,不同颜色代表不同的奖励大小(果汁滴数1-10)。中:训练集2,刺激所代表获得奖励的大小设为5固定不变,不同点数代表不同的获得奖励的概率(0.1-1.0)。下:实验设计包括6个不同条件,2(熟悉/陌生)ⅹ3(一致/单维/不一致)。(b)测试集:灰色格子代表训练过的熟悉选项,白色格子代表训练中未出现过的新选项,画叉代表该刺激不包括在测试集内;带数字的格子是6个不同条件的几个示例。(c)两个试次中刺激的示例。(d)实验1:决策任务和fMRI扫描;实验2:fMRI和单选项任务(无需决策);实验3:TUS干预和决策任务。(e)时间线:灰色代表采用训练集1和2隔日交替训练,红色代表实验1,蓝色代表实验2,黄色代表实验3。
在第一个实验中,猕猴需要在两种刺激之间做出决策,同时研究者对猕猴的大脑进行fMRI扫描。进一步对扫描得到的BOLD(blood oxygen level-dependent)信号通过广义线性模型(generalized linear model,GLM)回归,可以得到与价值比较相关的参数值。当动物做出更有利的选择(即实际的价值更高)时,该参数会得到正值,反之则为负值(无论正值或负值都能反映该区域能够表征价值比较的过程,符号反映的可能是更复杂的表征机制)。结果表明,不管刺激是熟悉的还是陌生的,在OFC(orbitofrontal cortex)和MFC(medial frontal cortex)两个脑区都能观察到对价值比较的表征;但OFC对熟悉和陌生刺激的反应无显著差异(Fig.2e-g),而前侧MFC则对熟悉刺激和陌生刺激的反应存在显著差异(Fig.2i-k)。第一部分实验结果表明OFC和MFC两个脑区都可能参与了价值编码的过程,但MFC可能在对陌生刺激的价值编码中有与众不同的作用。
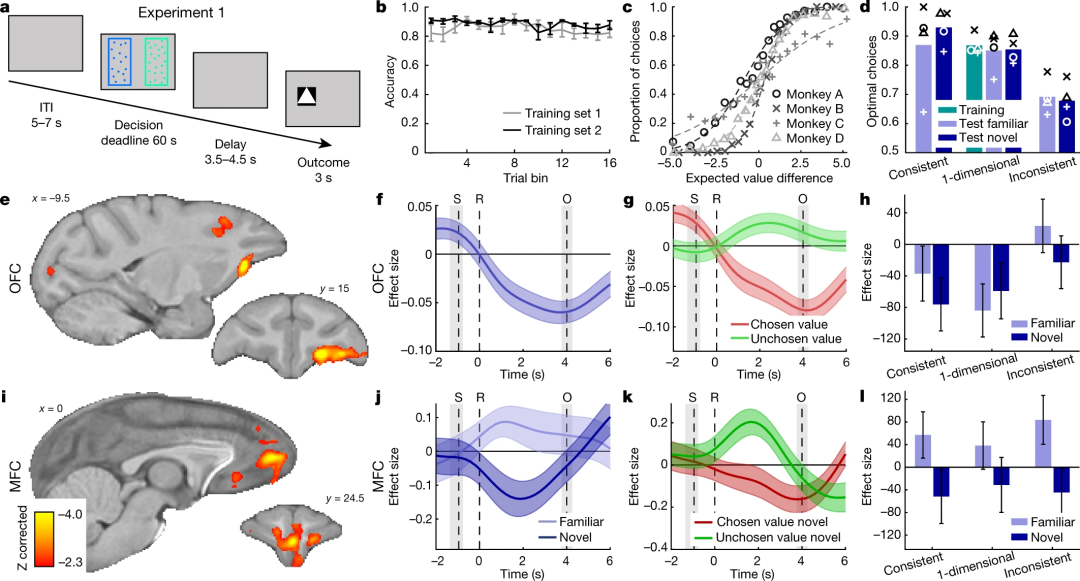
Fig.2|实验1,与熟悉/新的决策相关的神经活动
(a)单个试次的时间线,实验包括4只猕猴,每只动物完成12个实验单元。(b)训练中的反应正确率,数据点表示为均值和标准差。(c)测试表现,横轴代表左边选项价值减去右边选项价值,纵轴为选择左边选项的概率。(d)训练和测试表现的比较,柱状图代表了4只动物的平均结果。(e)和价值比较相关的信号。(f)OFC脑区BOLD信号效应量变化的时间曲线:S,刺激;R,反应;O,结果。(g-h)不同实验条件下的OFC价值比较信号:深紫色为新选项,浅紫色为熟悉选项。(i-l)MFC相关信号,意义同e-h
在第二个实验中每个试次中仅出现一个刺激(如Fig.3a,猕猴不需要做决策)。这一部分实验研究的变量是前后两个试次的刺激价值之间的关系。当两次呈现同一种刺激时,fMRI扫描得到的BOLD信号会在第二次减弱,即“重复抑制”(repetition inhibition)现象(反之,前后呈现不同刺激则不会出现此现象,如Fig.3b)。在OFC,研究者发现了很强的重复抑制现象;而在MFC却没有发现这种现象(Fig.3e)。这意味着OFC对价值的编码是基于特定选项的,而MFC对价值的编码则另有乾坤。
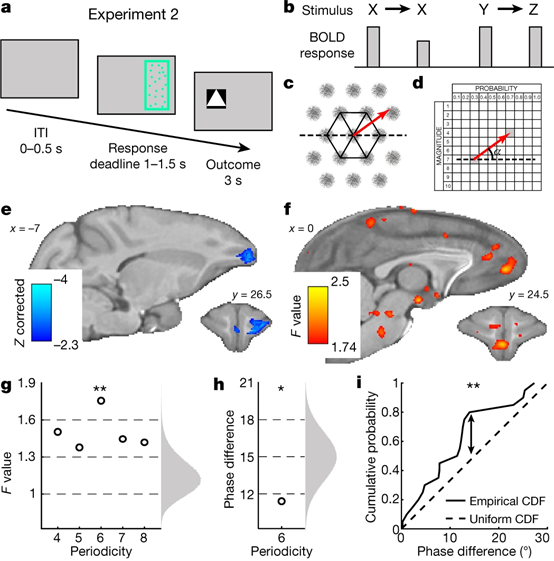
Fig.3 | 实验2,刺激和价值空间的神经表征
(a)单个试次的时间线,实验包括4只动物,每只动物完成5个实验单元。(b)重复抑制效应的示意图。(c)一个理想的网格细胞(灰色)的六角对称感受野(黑色)以及在不同位置间的一条轨迹(红色)。(d)在价值空间内上一个选项到下一个相邻选项的轨迹(红色),角度是相对于一个固定方向(虚线)计算的夹角。(e)同一个选项的重复抑制效应对应的反应区域。(f)针对六角对称的正交测试对应的反应区域。(g)其它角度为周期的正交测试结果,360/angle = n fold,例如 360/60= 6 fold。(h)同一个实验单元内将数据分半检验,价值空间内的网格的方向是不变的。(i)在不同实验单元内网格的方向有差异。
在人类被试上进行的fMRI扫描实验中,前人报告了当物理空间中的位置轨迹与六边形网格平行/不平行的条件下,内嗅皮层神经元的反应不同,揭示了内嗅皮层的神经元对物理空间的位置编码是依赖于一种独特的六角对称感受野(Fig.3c);因此Bongioanni等人猜测,在概念空间中,包括我们现在所讨论的价值空间,神经元的编码方式可能也是类似的。
Fig.3c中的红色轨迹代表着物理空间中的位置轨迹,类似地,在当前的研究中,猕猴就好比在一个价值空间内漫游,Fig.3d中红色箭头表示了实验中前后两个相邻的试次中的刺激的价值在价值空间形成的一条轨迹。研究者进一步发现,这条轨迹的角度(如Fig.3d中所示可定义为与价值空间中某一个固定方向的夹角)每转过60°,MFC神经元的活动也会随之发生周期性变化,而在OFC则未观察到相同现象(Fig.3f);并且以其它的角度转动也没有类似现象出现(Fig.3g)。此外,在同一个实验单元内将数据进行分半检验,结果表明价值空间内的网格的方向是不变的;但在不同实验单元之间,网格的方向有差异,这说明了价值空间内的网格可以重构映射(Fig.3h&3i)。
结合两个实验的结果,也就是说OFC和MFC对价值选择的过程均有表征,但背后的机制不同:OFC神经元是基于特定的选项进行价值编码,而MFC则在价值空间内以六边形网格的形式编码,后者使得猕猴能够整合多维度价值信息来评估新刺激的价值。

Fig.4|神经刺激对行为的影响
(a)TUS和任务时间线,实验包括3只猕猴,每个条件下完成4个实验单元,所有试次都是刺激的两个维度价值指向不一致的实验条件。(b)主观判断价值的模型:左,采用更优化的乘积方式(维度1×维度2);右,更简单的加法方式(维度1+维度2)/2。(c)利用模型计算的整合系数。(d)MFC的刺激位点和位于更后侧额叶的控制组刺激位点。(e)重复测量的方差分析结果,从左至右三组分别为:控制位点TUS刺激组,安慰剂组和MFC TUS刺激组。
对一个陌生刺激的编码,很重要的一点就是需要从中提取出经验中已有的与价值相关的特征。这些特征很有可能分布在多个维度上,因此需要神经元将多维度价值信息进行整合。整合的方式大致可能有三种:一种是根据两个维度价值的乘积进行整合,第二种是孤立两个维度,分别在两个维度评估价值后相加(Fig.4b所示为两种可能性对应的两个简化模型),第三种是两种策略的混合。研究者通过模拟实验,发现混合模型能更好地描述了真实数据。进一步地,研究者定义了一个“整合系数”(integration coefficient)来描述整合模型在混合模型中的权重。接着在第三个实验中,研究者采用干预手段,利用经颅超声刺激(transcranial ultrasound stimulation,TUS)扰乱MFC活动(Fig.4a),进而探讨MFC活动和对陌生刺激的价值编码之间的因果关系。研究者发现当采用TUS扰乱MFC后,整合系数显著地降低了,也就表明MFC在整合多维度的价值编码中扮演着重要角色。
结合三个实验的结果,当前的研究报告了猕猴的OFC和MFC两个脑区都能够编码价值,但二者的编码方式不同:OFC基于特定对象进行价值编码,而MFC则是在价值空间内进行网格式编码。MFC的编码特征更有利于大脑对陌生刺激的价值编码,而fMRI实验结果也证明了MFC对熟悉刺激和陌生刺激的反应的确有显著差异;通过干预实验,研究者进一步证明了MFC的活动与多维度价值信息整合之间的因果关系。
为什么你这么争强好斗?
pSI神经元早已未卜先知
“好斗”是动物的天性,只有保持着这种“勇者无畏”的闯劲,动物们才得以获得充足的食物和繁衍后代。当然,在不同性别、年龄的动物大脑中,控制攻击行为的区域不尽相同,并且在不同的情况下(比如不同的基因型、繁殖状态和社会环境中),各类攻击行为也是由各自不同的神经环路精准调控着。不能否认的是,在多数攻击行为中,典型的“教科书式”的表现常常包括撕咬、追逐、大声恐吓对方等(如你有幸未能亲身经历过这一切,请参考“动物世界”)。这说明很有可能存在着一些攻击行为共享环路。同样有些病人会在缺乏外界威胁的情况下发起不可控制的不合常理的攻击行为,更能说明脑区中很有可能存在着一些神经环路可以编码一个普遍的攻击行为应答。
但事实真的是这样吗?如果真的存在着这种协同环路,那它又是如何眼观四路耳听八方,并指挥机体进攻的呢?近日,来自浙江大学脑科学与脑医学学院的虞教授和段院士实验室合作,通过电生理和钙成像等技术共同发现了小鼠后侧无名质(posterior Substantia Innominata,pSI)的神经元可以根据小鼠面临的情况危急程度呈现出不同程度的活性,并预测接下来会发生的攻击行为方式。
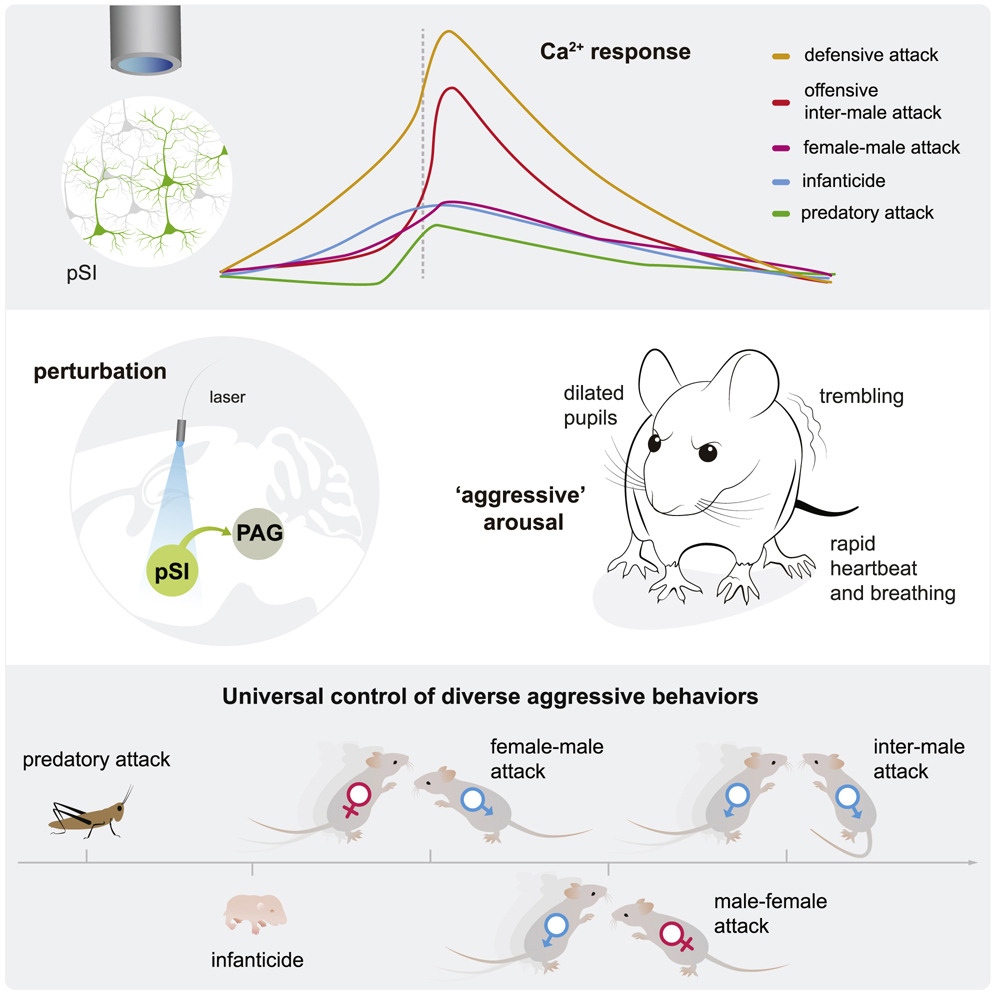 - Zhu et al。, Neuron。-
- Zhu et al。, Neuron。-因为无名质早在4年前就被曝出与攻击行为有关的情绪状态有关,所以,研究人员首先想知道,SI在攻击行为中到底起着什么作用?他们通过c-Fos染色观察到,小鼠处于雄性攻击状态中激活的pSI神经元数量远远高于它们处于探索的状态,并且通过记录pSI神经元的激活率确定了攻击状态中pSI神经元的活性更高。即使处于攻击前的“闻香识鼠”鉴别同类雄性的阶段,pSI神经元就已经嗅到了一丝不安,开始增强自身的活性,为一触即发的大战做好准备。所以说,pSI神经元在雄性攻击状态中是高度活跃的,那么它们极有可能可以编码诱发攻击行为的信号。接着,通过钙成像技术,研究人员更进一步分析了能表达Thy1的pSI神经元在雄性攻击状态不同阶段的活动,他们注意到pSIThy1神经元在攻击发生前就早早地开始活跃起来,相比于并未发生攻击行为前的“望闻问切”阶段,实际发生攻击前的同一时期pSIThy1神经元的钙活动更为活跃,也就是说,pSIThy1神经元的活动可以准确的预测接下来是否会启动攻击。
那么,面对环境中各式各样的信息,pSIThy1神经元是否能准确判断哪些是威胁、哪些是正常社交,并对此做出适当的反应从而调节动物的行为呢?研究人员发现,各种不同的威胁都会造成pSIThy1神经元强烈的应答,并且有趣的是,面对不同的威胁状况,pSIThy1神经元的活性成等级分布,威胁越强,神经元活性也越强,所对应接下来的攻击行为也更为激烈。pSIThy1神经元中的钙信号强度及持续时间整体可以准确地反映小鼠所面对威胁刺激的强弱:掠夺性攻击(predatory aggression)<攻击幼崽(pup-directed aggression)<攻击雌性(female aggression)<雄性进攻性攻击(male offensive aggression )<雄性防守性攻击(male defensive aggression)。
那激活pSIThy1神经元是否足以改变小鼠的攻击行为呢?研究人员激活pSIThy1神经元或激活下游的腹侧部中脑导水管周围灰质(ventral lateral/lateral PAG,VL/LPAG)后发现,小鼠的攻击性明显增强。更甚者,光激活这些投射到PAG的pSI神经元(pSI-PAG)的瞬间(约光照3s后),原本温和的小鼠武魂觉醒(呼吸加促、心跳加快和身体颤抖)并表现出很强的攻击性,立即对笼子里的对手发起猛烈的攻击。这个对手此时想必是一脸黑线吧,相煎何太急呢?研究人员认为这种光激活pSI-PAG神经元带来的武魂觉醒完全可以克服小鼠自身或者外界环境的影响,一手遮天。所以他们接着研究了在不同环境情况下,激活pSI-PAG神经元所带来的攻击行为变化。不论是独居或是群居或是处于陌生环境等不同的境遇,小鼠都会表现很强的攻击性,即使是昔日情人也难逃魔爪。
更为有趣的是,如果用不同光强造成不同程度的pSI-PAG神经元激活,带来的攻击行为结果也会随之改变,比如低刺激只会激发小鼠产生攻击幼崽和掠夺性攻击等较弱的攻击行为。同时他们注意到,如果小鼠刚刚经历了社会挫败,则引起它们表现相同攻击行为的神经元激活阈值则会升高,说明社会压力也可以调控pSI-PAG神经元。最后,研究人员也验证了pSI-PAG神经元只与攻击行为息息相关,对于交配等中立的行为则并没有太多的影响。
总的来说,这项研究揭示了小鼠大脑后侧无名质的神经元可以呈梯度编码预告多种攻击行为,可能解释了部分病态暴力行为的神经机制,为未来减少暴力的发生提出了新的解决方案。所以,每当你面对挑衅,pSI神经元都已经在暗暗地唤醒你的武魂,在你心跳加速,呼吸急促的同时怂恿着你用拳头解决掉对方。